



一、Повышение чувствительности реакционной системы:
1. Изолировать высококачественную РНК:
Успешный синтез кДНК происходит из высококачественной РНК.Высококачественная РНК должна быть как минимум полной длины и не содержать ингибиторов обратной транскриптазы, таких как ЭДТА или ДСН.Качество РНК определяет максимальное количество информации о последовательности, которую вы можете транскрибировать в кДНК.Обычный метод очистки РНК представляет собой одноэтапный метод с использованием изотиоцианата гуанидина / кислого фенола.Чтобы предотвратить загрязнение следовыми количествами РНКазы, РНК, выделенную из образцов, богатых РНКазой (таких как поджелудочная железа), необходимо хранить в формальдегиде, чтобы сохранить высококачественную РНК, особенно для длительного хранения.РНК, выделенная из печени крысы, в основном разлагалась после хранения в воде в течение одной недели, в то время как РНК, выделенная из селезенки крысы, оставалась стабильной после хранения в воде в течение 3 лет.Кроме того, транскрипты длиной более 4 т.п.н. более чувствительны к деградации следовыми РНКазами, чем небольшие транскрипты.Для повышения стабильности хранящихся образцов РНК РНК можно растворить в деионизированном формамиде и хранить при температуре -70°C.Формамид, используемый для сохранения РНК, не должен содержать обломков, разрушающих РНК.РНК из поджелудочной железы может храниться в формамиде не менее одного года.При подготовке к использованию РНК можно использовать следующий метод осаждения РНК: добавить NaCl до 0,2 М и 4-кратного объема этанола, поместить при комнатной температуре на 3-5 минут и центрифугировать при 10000×g в течение 5 минут.
2. Используйте обратную транскриптазу RNaseH-неактивную (RNaseH-):
Ингибиторы РНКазы часто добавляют в реакции обратной транскрипции, чтобы увеличить длину и выход синтеза кДНК.Ингибиторы РНКазы следует добавлять во время реакции синтеза первой цепи в присутствии буфера и восстанавливающего агента (такого как DTT), потому что процесс, предшествующий синтезу кДНК, денатурирует ингибитор, тем самым высвобождая связанную РНКазу, которая может расщеплять РНК.Ингибиторы белковой РНКазы предотвращают только деградацию РНК РНКазой A, B, C и не предотвращают попадание РНКазы на кожу, поэтому будьте осторожны, чтобы не ввести РНКазу с пальцев, несмотря на использование этих ингибиторов.
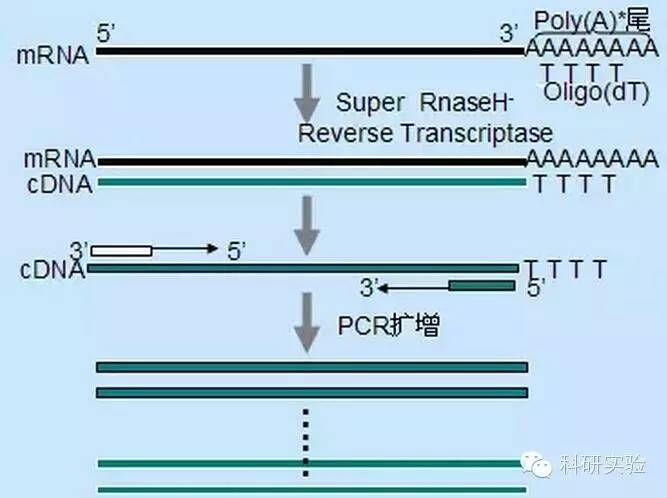
Обратная транскриптаза катализирует превращение РНК в кДНК.Как M-MLV, так и AMV обладают эндогенной активностью РНКазы H в дополнение к собственной полимеразной активности.Активность РНКазы Н и активность полимеразы конкурируют друг с другом за гибридную цепь, образованную между матрицей РНК и праймером ДНК или удлиняющей цепью кДНК, и разрушают цепь РНК в комплексе РНК:ДНК.Матрица РНК, деградированная активностью РНКазы Н, больше не может служить эффективным субстратом для синтеза кДНК, что снижает выход и продолжительность синтеза кДНК.Следовательно, было бы полезно устранить или значительно снизить активность РНКазы H обратной транскриптазы.。
Обратная транскриптаза SuperScript Ⅱ, обратная транскриптаза RNaseH-MMLV и обратная транскриптаза thermoScript, RNaseH-AMV, могут получать большее количество и более полноразмерную кДНК, чем MMLV и AMV.На чувствительность ОТ-ПЦР будет влиять количество синтезируемой кДНК.ThermoScript гораздо более чувствителен, чем AMV.Размер продуктов ОТ-ПЦР ограничен способностью обратной транскриптазы синтезировать кДНК, особенно при клонировании более крупных кДНК.По сравнению с MMLV, SuperScripⅡ значительно увеличивает выход продуктов длинной ОТ-ПЦР.РНКаза-Н-обратная транскриптаза также обладает повышенной термостабильностью, поэтому реакцию можно проводить при температурах выше обычных 37-42°С.В предлагаемых условиях синтеза используйте олиго(dT) праймер и 10 мкКи [α-P]dCTP.Общий выход первой цепи рассчитывали с использованием метода осаждения ТХУ.Полноразмерную кДНК анализировали с использованием отсортированных по размеру полос, вырезанных и подсчитанных на щелочном агарозном геле.
3. Поднимите температуру инкубации для обратной транскрипции:
Более высокая температура инкубации способствует раскрытию вторичной структуры РНК, увеличивая выход реакции.Для большинства матриц РНК инкубация РНК и праймеров при 65°C без буфера или соли с последующим быстрым охлаждением на льду устраняет большинство вторичных структур и позволяет праймерам связываться.Однако некоторые шаблоны все еще имеют вторичные структуры даже после тепловой денатурации.Амплификация этих сложных матриц может быть выполнена с использованием обратной транскриптазы ThermoScript и помещением реакции обратной транскрипции при более высокой температуре для улучшения амплификации.Более высокие температуры инкубации также могут повысить специфичность, особенно когда для синтеза кДНК используются геноспецифические праймеры (GSP) (см. главу 3).При использовании GSP убедитесь, что Tm праймеров совпадает с ожидаемой температурой инкубации.Не используйте oligo(dT) и случайные праймеры при температуре выше 60°C.Случайные праймеры требуют инкубации при 25°C в течение 10 минут перед повышением температуры до 60°C.В дополнение к использованию более высокой температуры обратной транскрипции, специфичность также может быть улучшена путем прямого переноса смеси РНК/праймеров с температуры денатурации 65°C на температуру инкубации обратной транскрипции и добавления предварительно нагретой двукратной реакционной смеси (синтез кДНК с горячим стартом).Этот подход помогает предотвратить межмолекулярное спаривание оснований, которое происходит при более низких температурах.Многократное переключение температуры, необходимое для ОТ-ПЦР, можно упростить с помощью термоциклера.
Термостабильная полимераза Tth действует как ДНК-полимераза в присутствии Mg2+ и как РНК-полимераза в присутствии Mn2+.Его можно поддерживать в тепле при максимальной температуре 65°C.Однако присутствие Mn2+ во время ПЦР снижает точность, что делает полимеразу Tth менее пригодной для высокоточной амплификации, такой как клонирование кДНК.Кроме того, Tth обладает низкой эффективностью обратной транскрипции, что снижает чувствительность, а поскольку обратную транскрипцию и ПЦР можно проводить с помощью одного фермента, контрольные реакции без обратной транскрипции нельзя использовать для сравнения продуктов амплификации кДНК с загрязняющей геномной ДНК.Продукты амплификации разделяли.
4. Добавки, способствующие обратной транскрипции:
Добавки, включая глицерин и ДМСО, добавляют в реакцию синтеза первой цепи, что может снизить стабильность двухцепочечной нуклеиновой кислоты и развязать вторичную структуру РНК.Можно добавить до 20 % глицерина или 10 % ДМСО, не влияя на активность SuperScript II или MMLV.AMV также может переносить до 20% глицерина без потери активности.Чтобы максимизировать чувствительность ОТ-ПЦР в реакции обратной транскрипции SuperScript II, можно добавить 10% глицерин и инкубировать при 45°C.Если в ПЦР добавить 1/10 продукта реакции обратной транскрипции, то концентрация глицерина в реакции амплификации составит 0,4%, что недостаточно для ингибирования ПЦР.
5. Лечение РНКазы H:
Обработка реакций синтеза кДНК РНКазой Н перед ПЦР может повысить чувствительность.Считается, что для некоторых матриц РНК в реакции синтеза кДНК предотвращает связывание продуктов амплификации, и в этом случае обработка РНКазой H может повысить чувствительность.Как правило, обработка РНКазой Н необходима при амплификации более длинных полноразмерных кДНК-мишеней-матриц, таких как малокопийный туберозный схероз II.Для этой сложной матрицы обработка РНКазой H усиливала сигнал, продуцируемый SuperScript II или кДНК, синтезированной AMV.Для большинства реакций ОТ-ПЦР обработка РНКазой Н необязательна, поскольку стадия денатурации ПЦР при 95°C обычно гидролизует РНК в комплексе РНК:ДНК.
6. Усовершенствование метода обнаружения малых РНК:
ОТ-ПЦР особенно сложна, когда доступны только небольшие количества РНК.Гликоген, добавленный в качестве носителя во время выделения РНК, помогает увеличить выход небольших образцов.Гликоген без РНКаз можно добавлять одновременно с добавлением тризола.Гликоген растворим в воде и может удерживаться в водной фазе с РНК для облегчения последующего осаждения.Для образцов менее 50 мг ткани или 106 культивируемых клеток рекомендуемая концентрация гликогена без РНКазы составляет 250 мкг/мл.
Добавление ацетилированного БСА в реакцию обратной транскрипции с использованием SuperScript II может повысить чувствительность, а для небольших количеств РНК уменьшение количества SuperScript II и добавление 40 единиц ингибитора нуклеазы RNaseOut может повысить уровень обнаружения.Если в процессе выделения РНК используется гликоген, все равно рекомендуется добавлять БСА или ингибитор РНКазы при использовании SuperScript II для реакции обратной транскрипции.
二、Повышение специфичности ОТ-ПЦР
1. Асинтез CND:
Синтез кДНК первой цепи можно инициировать тремя различными способами, относительная специфичность которых влияет на количество и тип синтезируемой кДНК.
Метод случайных праймеров был наименее специфичным из трех методов.Праймеры отжигаются во множестве сайтов по всему транскрипту, генерируя короткие кДНК неполной длины.Этот метод часто используется для получения 5'-концевых последовательностей и получения кДНК из РНК-матриц с участками вторичной структуры или с сайтами терминации, которые не могут быть реплицированы обратной транскриптазой.Чтобы получить самую длинную кДНК, необходимо эмпирически определить соотношение праймеров и РНК в каждом образце РНК.Начальная концентрация случайных праймеров составляла от 50 до 250 нг на 20 мкл реакционной смеси.Поскольку кДНК, синтезируемая из тотальной РНК с использованием случайных праймеров, представляет собой преимущественно рибосомную РНК, в качестве матрицы обычно выбирают поли(А)+РНК.
Праймеры Oligo(dT) более специфичны, чем случайные праймеры.Он гибридизуется с поли(А)-хвостом, обнаруженным на 3'-конце большинства эукариотических мРНК.Поскольку поли(А)+ РНК составляет примерно от 1% до 2% от общей РНК, количество и сложность кДНК намного меньше, чем со случайными праймерами.Из-за своей высокой специфичности oligo(dT) обычно не требует оптимизации соотношения РНК и праймеров и отбора поли(А)+.Рекомендуется использовать 0,5 мкг олиго(дТ) на 20 мкл реакционной системы.oligo(dT)12-18 подходит для большинства ОТ-ПЦР.Система ThermoScript RT-PCR предлагает oligo(dT)20 из-за его лучшей термостабильности при более высоких температурах инкубации.
Ген-специфические праймеры (GSP) являются наиболее специфичными праймерами для стадии обратной транскрипции.GSP представляет собой антисмысловой олигонуклеотид, который может специфически гибридизоваться с целевой последовательностью РНК, в отличие от случайных праймеров или олиго(dT), которые отжигаются со всеми РНК.Те же самые правила, которые используются для создания праймеров для ПЦР, применимы к дизайну GSP в реакциях обратной транскрипции.GSP может иметь ту же последовательность, что и праймер для амплификации, который отжигается с самым 3'-концом мРНК, или GSP может быть разработан для отжига ниже обратного праймера для амплификации.Для некоторых амплифицированных субъектов необходимо разработать более одного антисмыслового праймера для успешной ОТ-ПЦР, поскольку вторичная структура РНК-мишени может препятствовать связыванию праймера.Рекомендуется использовать 1 пмоль антисмыслового GSP в 20 мкл реакции синтеза первой цепи.
2. Поднимите температуру инкубации для обратной транскрипции:
Чтобы в полной мере воспользоваться всеми преимуществами специфичности GSP, следует использовать обратную транскриптазу с более высокой термостабильностью.Термостабильные обратные транскриптазы можно инкубировать при более высоких температурах, чтобы увеличить жесткость реакции.Например, если GSP подвергается отжигу при 55°C, специфичность GSP не будет полностью использована, если для обратной транскрипции используется AMV или M-MLV при низкой строгости 37°C.Тем не менее, SuperScript II и ThermoScript могут реагировать при температуре 50°C и выше, что позволит устранить неспецифические продукты, образующиеся при более низких температурах.Для максимальной специфичности смесь РНК/праймер можно перенести непосредственно из температуры денатурации 65°C в температуру инкубации с обратной транскрипцией и добавить в предварительно нагретую двукратную реакционную смесь (горячий старт синтеза кДНК).Это помогает предотвратить межмолекулярное спаривание оснований при низких температурах.Множественные температурные переходы, необходимые для ОТ-ПЦР, можно упростить с помощью термоциклера.
3. Уменьшает загрязнение геномной ДНК:
Потенциальная трудность, возникающая при ОТ-ПЦР, заключается в загрязнении геномной ДНК РНК.Использование хорошего метода выделения РНК, такого как реагент Trizol, уменьшит количество геномной ДНК, загрязняющей препарат РНК.Чтобы избежать продуктов, полученных из геномной ДНК, РНК можно обработать ДНКазой I уровня амплификации для удаления загрязняющей ДНК перед обратной транскрипцией.Расщепление ДНКазой I прекращали путем инкубации образцов в 2,0 мМ ЭДТА в течение 10 минут при 65°С.ЭДТА может хелатировать ионы магния, предотвращая зависимый от ионов магния гидролиз РНК при высоких температурах.
Чтобы отделить амплифицированную кДНК от загрязняющих продуктов амплификации геномной ДНК, можно сконструировать праймеры, каждый из которых отжигает отдельные экзоны.Продукты ПЦР, полученные из кДНК, будут короче, чем продукты, полученные из загрязненной геномной ДНК.Кроме того, на каждой матрице РНК проводили контрольный эксперимент без обратной транскрипции, чтобы определить, происходит ли данный фрагмент из геномной ДНК или кДНК.Продукт ПЦР, полученный без обратной транскрипции, происходит из генома.
Время публикации: 16 мая 2023 г.



